Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid
This paper, originally published in Nature in in 1953, unraveled the structure of DNA. This was such a huge contribution to biology that it won the authors Nobel Prize just 9 years after the publication. It’s amazing how DNA has become such a commonplace knowledge in less than 50 years. The paper is surprisingly readable, short and to the point.
Yellow highlights/annotations are my own. You can disable them.We wish to suggest a structure for the salt of deoxyribose nucleic acid (D.N.A.). This structure has novel features which are of considerable biological interest.
A structure for nucleic acid has already been proposed by Pauling and Corey[1]. They kind made their manuscript available to us in advance of publication. Their model consists of three intertwined chains, with the phosphates near the fibre axis, and the bases on the outside. In our opinion, this structure is unsatisfactory for two reasons:
- We believe that the material which gives the X-ray diagrams is the salt, not the free acid. Without the acidic hydrogen atoms it is not clear what forces hold the structure together, especially as the negatively charged phosphates near the axis will repel each other.
- Some of the van der Waals distances appear to be too small.
Another three-chain structure has also been suggested by Fraser (in the press). In his model the phosphates are on the outside and the bases on the inside, linked together by hydrogen bonds. This structure as described is rather ill-defined, and for this reason we shall not comment on it.
This figure is purely diagrammatic. The two ribbons symbolize the two phosphate-sugar chains and the horizontal rods the pairs of bases holding the chains together. The verticle line marks the fibre axis.
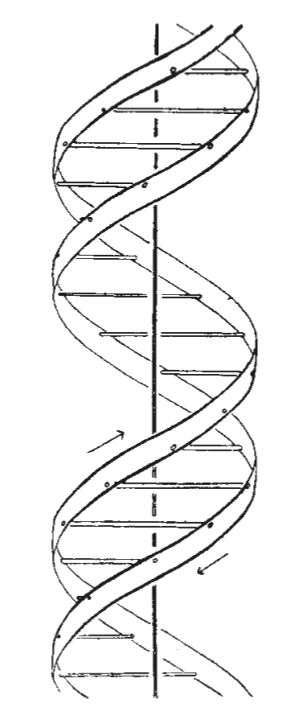
We wish to put forward a radically different structure for the salt of deoxyribose nucleic acid. This structure has two helical chains each coiled round the same axis (see diagram). We have made the usual chemical assumptions, namely, that each chain consists of phosphate diester groups joining ß-D-deoxyribofuranose residues with 3’, 5’ linkages. The two chains (but not their bases) are related by a dyad perpendicular to the fibre axis. Both chains follow right-handed helices, but owing to dyad the sequences of the atoms in the two chains run in opposite directions. Each chain loosely resembles Furberg’s model No. 1; that is, the bases are on the inside of helix and the phosphates on the outside. The configuration of the sugar and the atoms near it is close to Furberg’s ‘standard configuration’, the sugar being roughly perpendicular to the attached base. There is a residue on each chain in every 3.4 A. in the z-direction. We have assumed an angle of 36° between adjacent residues in the same chain, so that the structure repeats after 10 residues on each chain, that is after 34 A. The distance of a phosphorus atom from the fibre axis is 10 A. As the phosphates are on the outside, cations have easy access to them.
The structure is an open one, and its water content is rather high. At lower water contents we would expect the bases to tilt so that the structure could become more compact.
The novel feature of the structure is the manner in the which the two chains are held together by the purine and pyrimidine bases. The planes of the bases are perpendicular to the fibre axis. They are joined together in pairs, a single base from one chain being hydrogen-bonded to a single base from the other chain, so that the two lie side by side with identical z-co-ordinates. One of the pair must be a purine and the other a pyrimidine for bonding to occur. The hydrogen bonds are made as follows: purine position 1 to pyrimidine position 1; purine position 6 to pyrimidine position 6.
If it is assumed that the bases only occur in the structure in the most plausible tautomeric forms (that is, with the keto rather than the enol configurations) it is found that only specific pairs of the bases can bond together. These pairs are adenine (purine) with thymine (pyrimidine) and guanine (purine) and cytosine (pyrimidine).
In other words, if an adenine forms one member of a pair, on either chain, then on these assumptions the other member must be thymine; similarly for guanine and cytosine. The sequence of bases on a single chain does not appear to be restricted in any way. However, if only specific pairs of bases can be formed, it follows that if the sequence of bases on one chain is given, then the sequence on the other chain is automatically determined.
It has been found experimentally [3, 4] that the ratio of the amounts of adenine to thymine, and the ratio of guanine to cytosine, are always very close to unity for deoxyribose nucleic acid.
It is probably impossible to build this structure with a ribose sugar in place of the deoxyribose, as the extra oxygen atom would make too close a van der Waals contact.
The previously published X-ray data [5, 6] on deoxyribose nucleic acid are insufficient for a rigorous test of our structure. So far as we can tell, it is roughly compatible with the experimental data, but it must be regarded as unproved until it has been checked against more exact results. Some of these are given in the following communications. We were not aware of the details of the results presented there when we devised our structure, which rests mainly though not entirely on published experimental data and stereochemical arguments.
It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.
Full details of the structure, including the conditions assumed in building it, together with a set of co-ordinates for the atoms, will be published elsewhere.
We are much indebted to Dr. Jerry Donohue for constant advice and criticism, especially on interatomic distances. We have also been stimulated by a knowledge of the general nature of the unpublished experimental results and ideas of Dr. M.H.F. Wilkins, Dr. R.E. Franklin and their co-workers at King’s college, London. One of us (J.D.W) has been aided by a fellowship from the National Foundation for Infantile Paralysis.
References:
- Pauling, L., and Corey, R. B., Nature, 171, 346 (1953); Proc. U.S. Nat. Acad. Sci., 39, 84 (1953).
- Furberg, S., Acta Chem. Scand., 6, 634 (1952).
- Chargaff, E., for references see Zamenhof, S., Brawerman, G., and Chargaff, E., Biochim. et Biophys. Acta, 9, 402 (1952).
- Wyatt, G. R., J. Gen. Physiol., 36, 201 (1952).
- Astbury, W. T., Symp. Soc. Exp. Boil. 1, Nucleic Acid, 66 (Camb. Univ. Press, 1947).
- Wilkins, M. H. F., and Randall, J. T., Biochim. et Biophys. Acta, 10, 192 (1953).